
8. Razmnožavanje bakterija i kriva razmnožavanja
Rast je ravnomerno povećavanje svih delova bakterije. Posledica rasta je razmnožavanje. Brzina razmnožavanja je vreme koje protekne da se jedna bakterija podeli u dve, naziva se još i vreme generacije.
Bakterije se najčešće dele na nekoliko načina:
- binarna deoba gde od jedne nastanu dve nove jedinke,
- pupljenje stvara se pupoljak,
- grananjem,
- razmnožavanje L-oblika (ovi oblici imaju veliko telo sa jedinkama i kad ono pukne oslobađaju se mlade jedinke).
Kriva razmnožavanja
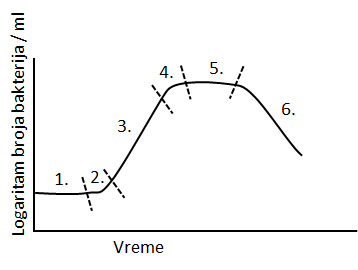
Foto Credit:Zona Medicine®
- faza pritajenosti – Lag faza je period prilagođavanja bakterija na novu sredinu, nema razmnožavanja.
- faza ubrzanog rasta – posle adaptacije bakterije se intezivno razmnožavaju.
- faza eksponencijalnog rasta – Log faza nastaje kada se uspostave optimalni uslovi.
- faza usporavanja – zbog povećanog broja bakterija dolazi do smanjene količine hranljivih sastojaka i nagomilavanja toksičnih produkata metabolizma. Dolazi do usporenog razmnožavanja i smrti dela populacije.
- stacionarna faza – rast je zaustavljen, bakterije su sitnije- broj umrlih i novostvorenih bakterija je jednak (ravna linija na krivulji).
- faza ubrzanog uginuća – nastaje kada se potroše rezerve hrane i kada se nagomilaju štetni produkti metabolizma. Bakterije umiru konstantno.
9. Faktori koji utiču na rast i razmnožavanje; kultivacija bakterija
Hrana obezbeđuje energiju za ćelije, ali treba da sadrži supstance koje ne stvara bakterija, kao što su faktori rasta (faktor V i X, itd.). Ako hrane ima više ili manje od optimalne količine dolazi do usporavanja rasta, razmnožavanja ili smrti.
Temperatura. Najbolja je optimalna. Prema temperaturi koju najbolje podnose, bakterije se dele na:
- psihrofilne bakterije. Vole niske temperature: Yersinia enterocolitica, Listeria…
- mezofilne bakterije. Vole sličnu temeraturu kao temperatura tela.
- termofilne. Vole visoke temperature.
Neke bakterije kao što su Campylobacter jejuni, coli rastu na 42oC, te ovaj podatak olakšava njihovu identifikaciju.
Koncentracija jona vodonika. pH koji odgovara najvećem broju bakterija je 5,5 do 8,5 pH. Neke poput Vibrio cholerae vole 8,5 pH. Dok lactobacilus voli nešto kiselije 5 pH.
Ugljen dioksid. Bakterije se dele na:
- autoorganotrofi koriste ovaj gas kao jedini izvor ugljenika
- heterotrofi ugljenik koriste iz organskih jedinjenja.
Kiseonik. U odnosu koju količinu kiseonika koriste bakterije se dele na:
- striktne anaerobe,
- aerotolerantne anaerobe,
- fakultativne anaerobe (mogu živeti i sa kiseonikom i bez njega),
- striktne aerobe,
- mikroaerofine bakterije.
Osim gore navedenih faktora važni su još i: vlaga, oksido-reduktivni potencijal, površinski napon, osmotski pritisak.
Kultivisanje bakterija
Postupak uzgajanja bakterija u veštačkoj sredini je kultivacija. Ona se sprovodi na veštačkim hranilištima. Hranljiva podloga treba da imitira prirodnu sredinu gde inače bakterija živi. Za autotrofne bakterije potrebne su obične podloge jer ugljenik dobijaju i ugljendioksida. Heterotrofne su značajne za humanu medicinu jer traže organski izvor ugljenika (glukozu na primer). Energiju dobijaju oksidacijom (aerobne bakt.) ili fermentacijom (anaerobne bakterije). Potrebe zavise od grupe bakterija. Escherichia coli koristi glukozu i druge šećere; Haemophilus influenzae faktore V i X. Hlamidije žive unutar ćelija, ne mogu se kultivisati na hranljivim podlogama. Bolesnički material se zasejava na veći broj hranljivih podloga. Inkubira se na određenoj temperaturi i trajanju od 24-36h.
10. metabolizam bakterija – Enzimi i ishrana
Sve hemijske reakcije koje se odvijaju u ćeliji bakterije nazivamo metabolizam. On podrazumeva dva osnovna procesa katabolizam i anabolizam. Katabolizam podrazumeva razlaganje složenih jedinjenja na prostija uz oslobađanje energije. Anabolizam je sinteza složenih jedinjenja od prostih uz utrošak energije. Karakteristike:
- metabolizam bakterija je do sto puta brži u odnosu na metabolizam eukariota,
- mogu da koriste različite supstrate kao izvor energije,
- sinteza složenih jedinjenja poput lipopolisaharida i teihoinske kiseline za bakterije je jednostavna.
Ishrana
Ishrana je najvažniji fiziološki proces. Hrana mikrobima služi kao izvor gradivnog materijala, kao izvor energije, sirovina za rezervne materije. Postoje dva tipa ishrane:
- holozoični – bakterije koriste čvrste čestice hrane, pa ih delovanjem svojih enzima razgrade, citoplazmatska membrane se uvija prema unutra (invaginacija) i unosi vanćelijsku materiju u ćeliju.
- holofitski – bakterije koriste u vodi rastvorljive hranljive materije koje prolaze preko celog ćelijskog zida i citoplazmatske membrane u obliku relativno malih molekula iz vodenog rastvora. Plazmalema je propustljiva za hranljive materije i vodu.
Prema potrebama za ugljenikom, bakterije delimo na:
- autotrofe – dobijaju ugljenik iz neorganskih izvora (CO2). Dobijaju azot iz neorganskih izvora (NH4, NO3). Neke bakterije ne mogu da vrše reakcije katabolizma bez ugljenika iz CO2: Brucella abortus, Neisseria gonorrhoea (treba im za rast); S. Aureus (za endotoksine); Bacillus anthracis (za kapsulu).
- heterotrofe – izvor ugljenika je organska materija. Azot iz organskih i neorganskih izvora.
Zavisno od potrebe za izvorom energije:
- fototrofi – koriste energiju elektromagnetnog zračenja za dobijanje nergije.
- hemotrofi – dobijaju energiju oksido-redukcionim reakcijama.
Zavisno od potrebe za donorom elektrona:
- litotrofi – donor elektrona je anorgansko jedinjejne,
- organotrofi – donor elektrona je organsko jedinjnje.
Enzimi
Enzimi su biološki katalizatori. Ubrzavaju hemijske reakcije nekoliko stotina puta. po hemijskoj strukturi su proteini. Učestvuju u metaboličkim reakcijama, disanju, pretvaranju i potrošnji energije, u sintezi makromolekula i ćelijskih komponenti. Enzimi povećavaju intenzitet hemijske reakcije, a pri tome se ne menjaju. Enzimi deluju na supstrate a supstance koje nastaju su produkti. Poznato je oko 2000 različitih enzima.
Struktura enzima:
Apoenzim je proteinski deo enzima, omogućava specifičnu adsorpciju molekula na enzim.
Prostetična grupa je neproteinski nedeljivi deo enzima.
Koenzim je organski kofaktor, vrši biokatalitičke enzimske reakcije, daje specifičnost enzimu, termostabilan je. Koenzimi mogu biti joni gvožđa, magnezijuma, vodonika. Važni koenzimi su NAD+ (nikotinamid adenin dinukleotid), NADP+ (nikotinamid adenin dinukleotin fosfat), FMN (flavin mononukleotid), FAD (flavin adenin dinukleotid), CoA (koenzim A).
Apoenzim + koenzim = holoenzim
Enzimi dobijaju naziv prema supstratu na koji deluje ili na osnovu njihovog zajedničkog delovanja (pr. Dehidrogenaze, karboksilaze). Nastavak na kraju označava cepanje (amilaza, želatinaza, katalaza, kazeinaza).
Bakterijski enzimi, podela prema prisutnosti u ćeliji:
- egzoenzimi. Izlučuju se u okolinu ćelije radi razgradnje velikih molekula na manje koje mogu ući u ćeliju difuzijom ili pomoću proteinskog nosača.
- endoenzimi. Katabolizuju biohemijske procese unutar bakterijske ćelije.
Bakterijski enzimi, podela prema supstratu na koje deluju:
- proteolitički,
- lipolitički,
- saharolitički.
Faktori koji utiču na aktivnost enzima su temperatura, pH i koncetracija supstrata. Regulacija sinteze se vrši kontrolom ekspresije bakterijskih gena. Regulacija aktivnosti već sintetizovanih bakterijskih enzima vrši se mehanizmom povratne sprege.
11. metabolizam energije; fermentacija; aerobna i anaerobna respiracija
Metabolizam energije
Energija je potrebna za transport jedinjenja u i iz ćelije, za kretanje i za biosintezu supstanci neophodnih za život. Bakterije energiju dobijaju oksido-reduktivnim procesima. Tokom reakcije oksidacije nekog jedinjenja dolazi do oslobađanja energije, i omogućava seodvijanje metaboličkih procesa koji troše tu energiju. Tokom oksido-redukcionih procesa troši se i stvara ATP.
Izvori energije
- Svetlost,
- neorganski izvori energije,
- organski izvori energije.
Fotoautotrofi. Koriste svetlost za stvaranje energije i ugljen-dioksid kao izvor ugljenika.
Hemoautotrofi. Koriste neorganska jedinjenja (H2S, NO3…) kao izvor energije, a ugljen-dioksid kao izvor ugljenika.
Hemoorganotrofi ili hemoheterotrofi. Koriste organska jedinjenja za energiju i ugljenik (lipidi, šećeri…). Bakterije iz ove grupe su najznačajnije za medicinu.
Fermentacija
Fermentacija je anaerobni proces za stvaranje energije. U ovom procesu glikolizom nastaje piruvat (2 ATP). On prima elektrone. Najveći broj bakterija koriste glikolizu za dobijanje piruvata.
Aerobna respiracija
Acetil CoA dobijen iz piruvata ulazi u Krebsov ciklus. Kiseonik je krajnji akceptor elektrona. Na kraju reakcije nastaje ugljen-dioksid i 34 molekula ATP.
Anaerobna respiracija
Krajni akceptor elektrona su sulafati, nitrati… Nastaje manje ATP, te su ove bakterije na nižem stupnju razvoja od onih koje koriste aerobnu respiraciju.
12. Bakterijski genom – hromozom bakterija
Bakterija nema jedarnu membranu, genetski materijal je okružen citoplazmom. Bakterija ima samo jedan hromozom – haploidna. Bakterija nema histone. Pored hromozomske DNK ima i plazmide. Bakterijski hromozom čini dvolančana DNK. Osnovna jedinica je nukleotid. Njega grade purinska ili pirimidinska baza (adenin, guanin; citozin, timin); molekul šećera pentoze i fosfatni radikal. Dva lanca su međudobno spojena vodoničnim vezama između odgovarajućih purinskih i pirimidinskih baza (A-T; G-C). DNK lanac je više puta izuvijan kako bi ceo stao u hromozom. Uloga DNK je:
- očuvanje identiteta vrste i
- determinacija strukturnih i funkcionalnih karakteristika.
Ovakvi procesi se odvijaju uz pomoć replikacije i transkripcije (sinteza RNK spram DNK).
13. Promenjivost bakterija – fenotipske i genotipski promene
Genom bakterije je zaštićen sistemom za ispravljanje grešaka, ali u njemu ipak dolazi do manjih ili većih promena. Osim toga bakterije manjaju svoje osobine i bez promena u DNK usled prilagođavanja na uslove u sredini u kojoj se nalaze. Pojava novih osobina koje su posledica promena u genomu, označava se kao genotipska promena. Adaptacija bakterije bez promena u genomu je fentotipska promena.
Fenotipske promene
Nastaju usled prilagođavanja bakterija bez promena u genotipu. Fenotipske promena su reverzibilne. Traju dok se uslovi životne sredine ne vrate u normalu. Primer:
- L-forma bakterija. Ove bakterije gube ćelijski zid u sredini gde je pencilin. Ali ako se premeste u sredinu bez antibiotika ćelijski zid se vraća;
- Kapsula S. pneumoniae. Kultivisanjem in vitro gube kapsulu, ali inokulacijom u miša geni za kapsulu se reaktiviraju;
- S i R varijacije. Isti soj bakterija može da ima glatke i rapave kolonije. Rapave nastaju kao posledica sinteze kapsulu, flagela…
Značaj fenotipskih promena se ogleda u ometanju delovanja antibiotika i otežanoj laboratorijskoj identifikaciji.
Genotipske promene
Genotipske mutacije su nasledne promene redosleda baza u lancu DNK. Mogu biti vidljive praćene fenotipskom ekspresijom, ili skrivene. Gen na kome nastaju promene zove se divlji alel, a ostali nastali njegovom mutacijom su mutantni aleli. Mutacije mogu biti prirodne i indukovane u laboratorji. Agensi koji izazivaju mutacije su:
- hemijski,
- fizički (toplota, UV, jonizujće zračenje…);
- biološki (segmenti DNK koji se ugrade u bakterijski genom).
Mutacije prema veličini mogu biti mikrolezije i makrolezije. Makro se dešavaju kao:
- delecije. Isecanje jednog dela lanca;
- duplikacije. Deo sekvence se dva puta ponavlja;
- inverzije. Rotacija segmenta DNK;
- insercije. Ugradnja segmenta DNK iz hromozoma, plazmida.
14. Genske rekombinacije; transformacija (Griffitov ogled)
Većina bakterije može da prenosi genetski materijal. Ćelija koja daje fragment DNK je donor, a ona koja prima je recipijent. Prenesena DNK može biti uništena dejstvom endonukleaza, te se genske rekombinacije najuspešnije obavljaju između srodnih bakterija. Postoje tri načina prenosa:
- transformacija,
- transdukcija,
- konjugacija.
Transformacija
Predstavlja prihvatanje DNK iz spoljašnje sredine i transport u unutrašnjost ćelije. Njuška koja je uočila ovaj način intime bakterija je Griffit. Ispitivao je karakteristike kapsule i njenog uticaja na virulentnost pneumokoka. Kako?
Ubrizgavao je virulentne i avirulentne (sa i bez kapsule) sojeve u eksperimentalnu životinju. Takođe je koristio toplotom ubijene virulentne pneumokoke, koje je ubrizgavao same i u smeši sa živim avirulentnim sojem pneumokoka. Zaključak:
- virulentni sojevi su ubijali miševe,
- avirulentni sojevi su ostavljali miševe u životu,
- toplotom ubijeni pneumokoki su takođe bili fini,
- smeša ubijenih virulentnih sa živim avirulentnim bila je letalna za miševe.
Grifit je zaključio da ubijene virulentne pneumokoke su na neki način prenele živim avirulentnim sposobnost sinteze kapsule.
15. Transdukcija i bakteriofagi
Transdukcija predstavlja prenos genetskog materijala iz donorske u recipijentnu ćeliju posredstvom bakteriofaga. Bakteriofagi su virusi koji imaju izražen afinitet za parazitiranje u bakterijskoj ćeliji. Postoji nekoliko grupa bakteriofaga A, B, C, D, E i F. Ovako raspoređeni bakteriofagi razlikuju se po spoljašnjem izgledu. Bakteriofag deluje tako što prepozna receptorsko mesto na ćeliji i vezuje se za njega uz pomoć fibrila. Ubrizgava svoj genom u bakterijsku ćeliju. Parazitira u njoj i koristi je kako bi se umnožio. Razlikuju se virulentni i umereni fagi. Virulentni proizvode veliki broj kopija i izazivaju lizu ćelije ili se postepeno oslobađaju procesom pupljenja.
Umereni fagi imaju sposobnost ugradnje u bakterijski DNK (profag), gde nastavljaju zajedno da se replikuju sa bakterijskim hromozomom. Profag može da pređe u litički ciklus razmnožavanja. Bakterijska ćelija sa inkorporiranim bakteriofagom označava se kao lizogena. U zavisnosti od mesta inkorporacija profaga mogu se izgubiti ili pojaviti nove osobine. Primeri:
Difterični efzotoksin sintetišu samo oni sojevi Corynebacterium-a diphtheriae koji su inficirani β-profagom; slično je dokazano i kod Clostridium botulinum-a (botulinski egzotoksin); i kod Sterptococcus pyogenes-a (pirogeni toksin). Ova pojava sticanja novih osobina bakterija u stanju lizogenije (u prisustvu određenog profaga) naziva se lizogena konverzija. Fag tokom litičnog ciklusa razmnožavanja formira određen broj nekompletnih kapsida i sadrži kopiju virusnog genoma. U nekompletni kapsid može upakovati fragment bakterijske DNK. To su zapravo transdukujići fagi. Transdukujići fag ima kompletan proteinski omotač. Kada napusti ćeliju domaćina može da prepozna receptore na drugoj bakterijskoj ćeliji, veže se i ubrizga svoju nukleinsku kiselinu. Ubrizgana nukleinska kiselina je bakterijskog porekla te neće doći do infekcije. Stvara se prilika za rekombinaciju donorske DNK (ubrizgane od strane bakteriofaga) sa hromozomom recipijentske ćelije. Ovaj proces prenosa DNK između dve ćelije naziva se generalizovana transdukcija.